What You Are

Some time ago, you were a single cell. Now, you’re ~35 trillion. Your trillions of cells all share the same DNA, half from mom and half from dad, yet perform functions as varied as perceiving music, digesting protein, and thinking. All in chaotic yet graceful coordination, to keep you alive.
In 1981, almost 30 years after Watson and Crick discovered the double helix structure of DNA, a Cambridge biologist named Rupert Sheldrake asked a question that all but got him exiled from mainstream science: How do cells coordinate to form exquisitely precise organs like hands, eyes, lungs, or brains? The short (but incomplete) answer is: DNA. The long answer remains a mystery, being worked on by two biologists whose work, in my opinion, validates Sheldrake’s pioneering search.
I want to tell you their story because it’s one of the most fascinating I’ve ever learned about, and because it’s literally the story of how you came to be as you are.
I. Morphogenesis
You are, in a very real sense, a bag of cells. All of your cells share a copy of the same DNA: your complete genome is inside every single one of them.
Twentieth-century biology was high on this discovery: DNA, the master blueprint of life. The source code. We had finally found the basis for Darwin’s theory of evolution: a programmable molecule that lets organisms adapt across generations.
But not everyone was so hyped.

Meet Sheldrake. To Sheldrake, DNA could not possibly be the complete explanation.
Yes, DNA explains what proteins a cell makes. It cannot, however, explain how cells carrying the same code know to become a liver cell here and a neuron there. How do multicellular organisms with differentiated organs arise from a single “source code”?
To think that DNA can explain why we have five fingers in each hand is like saying quantum mechanics can explain why Napoleon lost the Battle of Waterloo. Yeah, it technically does, but that’s not a sufficient explanation.
DNA brought cells into the realm of informatics; they’re quite literally self-programmable biochemistry. As an explanatory framework, it was outrageously prolific. Thanks to our understanding of DNA, we could decode the low-level language of life.
But the mechanics of multicellular life, both at the historical and developmental level, remain unbeaten by DNA: Exactly how did cells organize into cooperative clusters in evolutionary history? And how do cells of a growing body coordinate to know which should form each organ?
Life should be unicellular. The fact that bodies made of trillions of cells exist is a miracle. And it’s literally the story of how you came to be in the world, not a mere theoretical curiosity! When I understood the magnitude of this mystery, I was even more surprised that there’s no mainstream, unified explanation. But I did find some satisfying theories, and they all start with Sheldrake’s question:
How does a single cell become a body?
To poor old Sheldrake’s misfortune, however, he was asking this question during peak “genes explain everything” optimism. Contemporary scientists didn’t really understand what he was asking, in my opinion. He was shunned by mainstream science, and it took 40 more years and two new generations of subversive biologists willing to revisit his legacy and take on the biggest open question in biology:
How do we come to be what we are?
II. Morphic Resonance
Sheldrake’s own failed attempt to answer the question is interesting in its own right.
In a nutshell, he proposed that species have a memory that takes the form of a field. This field contains the body plan for a species. When a new organism is created after the gametes combine, the developing body somehow tunes in to this so-called morphic field and grows into the proper form. Sheldrake posited that this happens via resonance between organisms.
This might sound esoteric, but resonance is a well-studied phenomenon in acoustics, where a signal amplifies another by vibrating synchronously. He also drew inspiration from what happens with inorganic crystallisation. Once synthesized, crystals become easier to crystallize everywhere in the world.
So, Sheldrake reasoned, when a particular form arises in nature, it creates a “morphic field” that makes it easier for the same form to arise again, by resonating with fields in future developing organisms.
The theory was elegant and ambitious, but wrong. Sheldrake was correct in identifying a gap between genes and bodies. But, as often happens with pioneering ideas in science, he appealed to the metaphysical for the explanation. Sheldrake argued his morphic field was non-physical, an explanation that most of us won’t accept and, even worse, can’t be tested. He had the right intuitions, as we will see, but the wrong framework.
The clues, it turns out, were hiding in two places Sheldrake never looked: cell membranes and mitochondria.
III. Bioelectricity
All your cells have an electric charge, not just neurons.
Every cell’s membrane maintains a voltage difference between its inside and outside, its membrane potential. And cells can share their electrical states with their neighbors through tiny channels called gap junctions. This means that the tissues in your body are running an electrical network nonstop. A slower version of the same kind of signaling that powers the nervous system.
It’s clear that this electrical network, often called “bioelectricity”, carries some information. Michael Levin, computer scientist turned biologist, asked: What if this electrical network is related to morphogenesis?
He did fascinating experiments with planaria: flatworms that can regenerate. If you split a planaria in half, they re-grow into two individuals in a matter of days. Even if you cut one into twenty (!) pieces, you get twenty new clone worms. The genes, of course, don’t change. So what’s telling the cells at the wound site what to grow (e.g., a head and not a tail)?
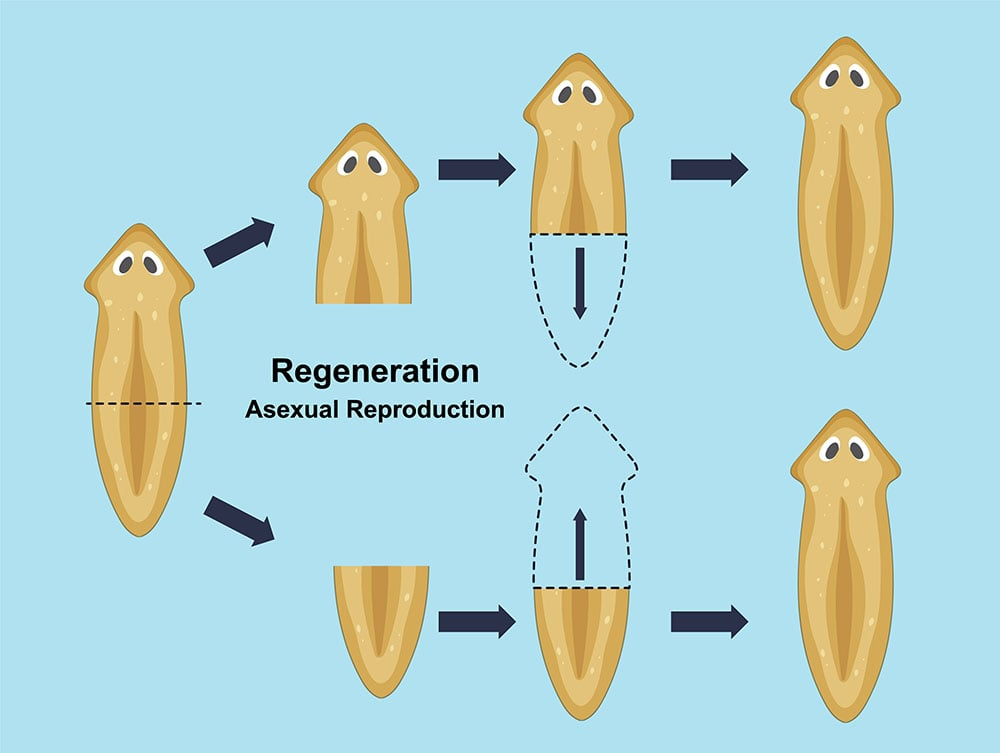
Levin found the answer was voltage. The cells in a planarian maintain a specific bioelectric pattern that encodes what belongs there. Membrane voltage functions as a kind of positioning system for where the cell is in the body. And when Levin’s team manipulated that pattern (by forcing cells to adopt different voltages via ion channel drugs), he could make a worm regenerate with two heads instead of one. Each head has its own brain and appears to have independent decision-making.
Moreover (!!!) when he cut those two-headed worms yet again, they kept regenerating as two-headed worms, on their own. The original genome was untouched: DNA was still normal planaria, and it encoded proteins for both the head and the tail (and everything else). But the bioelectric pattern had been overwritten, and the organism persisted the new two-headed one, creating a stable memory of the form it “should” have, which is clearly stored outside the genome. This experiment alone proves that genes are not sufficient (!) to explain an organism’s body plan.
Levin then moved to frogs. By injecting ion channels that shifted the voltage of cells in a developing frog’s gut, he “forced” those cells to create a fully functional eye. Let me repeat that: he made AN EYE GROW OUT OF A FRIGGIN TADPOLE’S GUT. Again, no DNA was modified. Only electric patterns between the growing cells. How did he do this? Levin studied the voltage pattern of naturally developing eyes (which is roughly 10 mV more negative than surrounding cells) and artificially reproduced it in another location.
Only a specific narrow window of membrane voltage produced eyes: if the voltage was too high or too low, nothing happened. But when the voltage was just right, it spawned the very complex genetic program for building a complete eye (lens, retina, etc.) without needing to micromanage genes or cell types. The misplaced eye is functional, btw, connects to the spinal cord on its own, and even allows the tadpole to see!
By now, I hope that any ideas you may have had about genes fully determining body shape were demolished. Remember, no DNA was modified in any of these experiments.
What does this mean for us, humans? It strongly suggests our bodies are determined by bioelectric patterns too. We have some indirect evidence: genetic disorders that disrupt ion channels cause morphological defects, and human cells in the lab respond to bioelectric manipulation. We don’t know for sure whether mom contributes to creating the right electric pattern, or whether the fetus creates it independently. My amateur armchair biologist guess is: it’s a little of both.
Electrical charge in cells determines which genes are expressed, then. Genes generate a field, and the field somehow determines which other genes get expressed. This creates a feedback loop, which Levin showed can be tinkered with.
Levin’s work is, in my opinion, groundbreaking. He shows bioelectricity is a computational layer that sits between the genome and the body plan. DNA specifies all the proteins a cell can make. But the bioelectric network that emerges carries its own information: a kind of map that tells cell collectives what shape to build, how big to grow, and when to stop. In a growing organism, DNA is “machine code”, and bioelectricity is like a UI (driven by DNA, but it can also modify its execution).
And, when you’re dealing with the right level of abstraction, you can hack things:
Michael Levin is a biohacker in the true sense of the word, and I predict he will win a Nobel Prize for his discoveries.
In a sense, Sheldrake was vindicated. He had predicted that a field beyond DNA encodes the body plan, that this field stores a memory of form, and that it can be rewritten. What Levin found is exactly that.
Sheldrake’s mistake was proposing a non-physical mechanism (morphic resonance) to explain the field. The real mechanism turned out to be bioelectricity: physical, measurable, and manipulable. But getting the mechanism wrong while getting the structure right is one of the most common patterns in the history of science. In my book, Sheldrake is a genius who was ahead of his time.
IV. The Origins of Multicellularity
Michael Levin’s work explains how cells coordinate to build a body plan. But there’s an even more basic question underneath: Why do cells coordinate at all? How did bioelectric networks that encode body plans arise, historically speaking?
For roughly two billion years, life on Earth was single-celled. Bacteria appeared as soon as the planet cooled, and then they kinda chilled. No hint of multicellularity, organs, or body plans. What was the bottleneck? Why did life on Earth stay at single-cell bacteria-level complexity for two billion years before “inventing” complex body plans? Why couldn’t bacteria build bodies?
Attempts to explain this informationally were always insufficient. Again, DNA was not the answer. Bacteria are genetic innovators: they swap DNA freely, evolve really fast, replicate ~hourly (i.e., 1 day = ~24 generations!), and have had billions of years and astronomical population sizes to explore every possible genetic trick. If the right genes were all it took, bacteria should have figured out multicellularity long ago.
So, it doesn’t seem to be an information optimization problem. What else can it be? Nick Lane’s answer: It was an energy issue. His book The Vital Question provides a deeply satisfying explanation.
Let’s try to reverse-engineer the problem by looking at existing multicellular-capable cells: eukaryotes.
We humans are made of what are called eukaryotic cells. All animals, plants, fungi, and anything with a body has eukaryotic cells only. These are very complex, very big cells, compared to our bacterial ancestors. For reference, a human skin cell is 1000x the size of an E. coli bacterium. Only this kind of huge cell seems to form organs and bodies. Why can’t bacteria?
The problem: bacteria produce energy (ATP) across their cell outer membrane. Their metabolism is based on pumping protons outside their limits, and having them flow back via a motor-like mechanism that synthesizes ATP (called, very creatively, ATP synthase). If they scale in size, volume grows with the cube, but membrane surface area only grows with the square. So a bigger bacterium actually gets worse energy per unit volume. How do eukaryotes solve this?
Enter the mitochondria!
Eukaryotes have what Nick Lane calls a chimeric origin: a cell got inside another cell, and the two began cooperating. The outer cell became eukaryotes, and the inner cell became mitochondria.
(If you’re like me, that sounds almost too good to be true. How likely is something like this to happen? Apparently, a cell entering another cell happens a lot. The hard part is having them cooperate. Think about it: both cells have developed their own genes for survival, treating everything else as a resource or a threat. One cell engulfing another, somehow not digesting it, somehow integrating two genomes, somehow coordinating two replication cycles. Seems like it shouldn’t be possible! But the evidence of this happening (at least once) is everywhere: all complex life! Nick Lane does a great job of reconstructing how such a miracle could come to be, and you should read his book if interested. Here’s my takeaway from his book: It probably almost happened millions of times and never worked, until one lineage stumbled into a stable arrangement (the inner cell lost the right amount of genes, a nucleus protected the outer cell’s genes, and sex was invented. For more details, read Lane’s book!))
All animals, plants, and fungi have mitochondria, and depend on them to generate ATP, life’s battery pack. Bacteria do not have mitochondria. By the way, most eukaryotes have thousands of mitochondria, not just one! Mitochondria is the plural of mitochondrion.
So, what do mitochondria do for our cells? Mitochondria decouple ATP generation from the membrane, solving the quadratic vs. cube energy per unit lockdown bacteria are trapped in. Eukaryotes broke this constraint by internalizing ATP production inside the cell, into mitochondria’s membranes, instead of the cell’s external membrane. Mitochondria are basically decentralized power plants floating around in our fat-ass eukaryotic cells. Actually, proton dams. Genius!
Unlocking (energy-efficient) bigger sizes enables eukaryotes to contain even more energy-churning mitochondria inside them, which in turn unlocks the next trick: huge genomes and lots of free energy available to “execute” them.
Another difference between bacteria and us is the size of our genome. Bacteria have ~1 megabyte genomes, while eukaryotes have ~1 gigabyte genomes. This means eukaryotes have much more code available to execute!
(btw: converting between base pairs and bytes is correct. Each base pair is one of 4 letters (A, T, G, C), so it takes exactly 2 bits to encode. That means 4 base pairs = 1 byte. For example: E. coli, the queen of bacteria, has ~4.6 million base pairs, equivalent to ~1.15 MB of code, while we humans have ~3.2 billion base pairs, equivalent to ~750 MB of code.)
Bacterial genomes are tiny compared to ours. A bacterium’s genome is a monstrously optimized piece of code. It needs to be, because they can’t afford the energy for big genomes! Why? Remember that RNA is self-executing code. But executing each gene requires a lot of energy! Carrying extra RNA costs energy to copy and maintain, so natural selection ruthlessly strips it out. Evolution strongly pressures bacteria to reduce their genome to the bare minimum.
We eukaryotes, with our neat distributed-energy-generation mitochondria trick, have ~200,000x more energy available per gene. This superabundance of ATP availability enables eukaryotic cells to afford to keep DNA around that doesn’t directly code for proteins. Some of that extra code becomes regulatory DNA: instructions that control when and where other genes get expressed (i.e., executed). Again, remember that RNA is basically bytecode, executable code. The larger programs you can store, the more logic you can try out and evolve.
That, Nick Lane believes, is how the jump from single-celled organisms to multicellularity happened. It’s a matter of energy availability. Bacteria were energy-starved and could only afford tiny genomes for each cell. Once the key innovation of mitochondria occurred, brought by a rare endosymbiosis event two billion years ago, cells had so much energy that they could push life to the complexity we know today: sexual reproduction, massive coordination (multicellularity), specialized tissue formation (organs), cell nucleus, adhesion molecules, programmed cell death, etc.
Lane answers the evolutionary half of Sheldrake’s question. How did cells organize into cooperative multicellular clusters in evolutionary history? By generating a superabundance of energy after the happy accident of engulfing another cell that turned into mitochondria.
V. Assembly
So let’s put it together.
You started as a single cell.
That cell had some pretty powerful tools.
- Mitochondria, inherited from mom.
- A genome, inherited from mom and dad.
Armed with this toolkit, you entered the world.
Your first cell divided, and divided again, and again, and each daughter cell inherited that same toolkit. As the cluster grew, a voltage pattern emerged across the cell collective: bioelectricity. That pattern told each cell where it was and what it should become, activating specific genes while silencing others. And the mitochondria, thousands per cell, provided the energy to actually run those massive genetic programs. From one cell, nurtured by mom, came your full body.
So, back to our original question: what are you?
You are an unbroken chain of organic biochemistry who found entropy pockets to exploit.
A four-billion-year legacy of matter becoming aware of itself.
A weird vehicle your selfish genes built to make copies of themselves.
A bag of cells, which in turn are a bag of mitochondria, ever-churning energy machines.
An electric field that reconfigures matter around it to produce a persistent and self-sustaining form.
A physical configuration of atoms that somehow resists the universal tendency to decay that is stipulated by the second law of thermodynamics.
The human body achieves tight coordination of trillions of autonomous agents (your cells) into a unified system. Multicellular bodies truly are the most impressive engineering achievement in the known universe. It was made possible thanks to the thousands of mitochondria in each of your cells, providing abundant energy to execute a body plan coordinated via bioelectricity.
That, little one, is what you are. Welcome home!

If you liked what you read, please consider subscribing below.
Further Reading
- Developmental bioelectricity wikipedia page
- “A revolution in biology”, article summarizing Levin’s work
- “The Vital Question” by Nick Lane
- Lex Fridman interviews Nick Lane
- Lex Fridman interviews Michael Levin
- Nick Lane: Origin of the eukaryotic cell
- My, Your Eyes Are So Electric
Acknowledgments
Cover photo by David Goodsell.